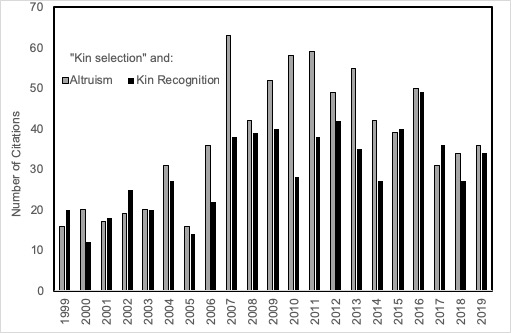
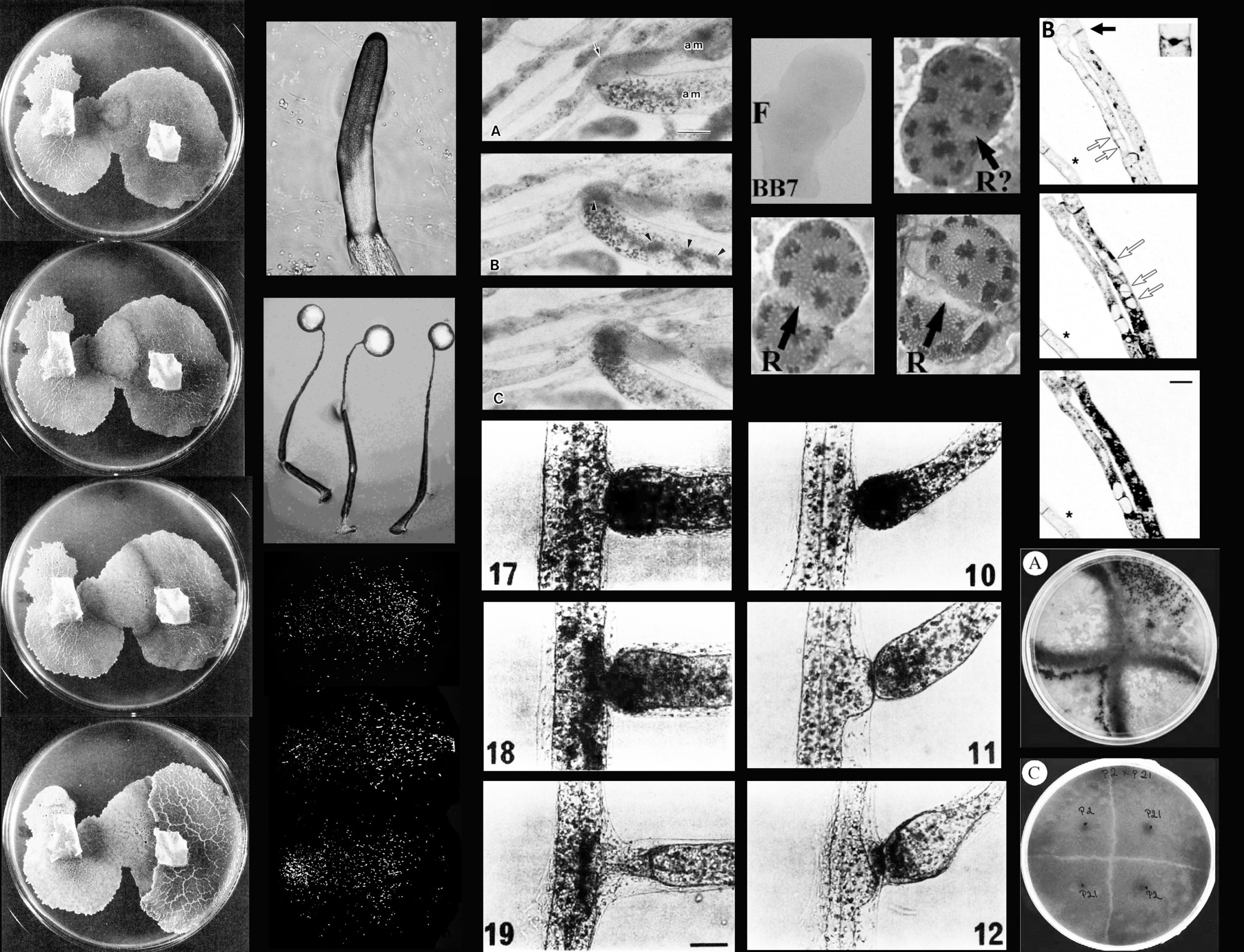
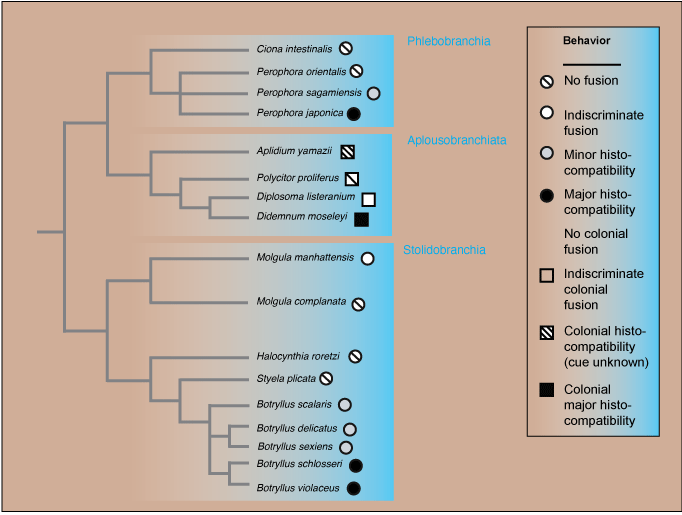
Resolution of anomalies
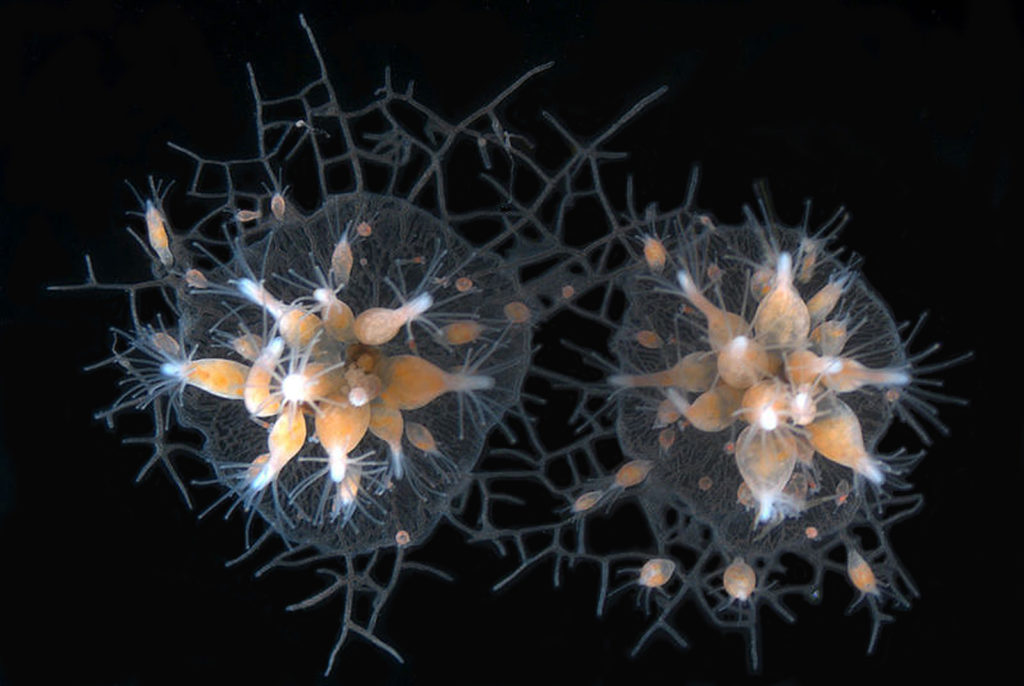
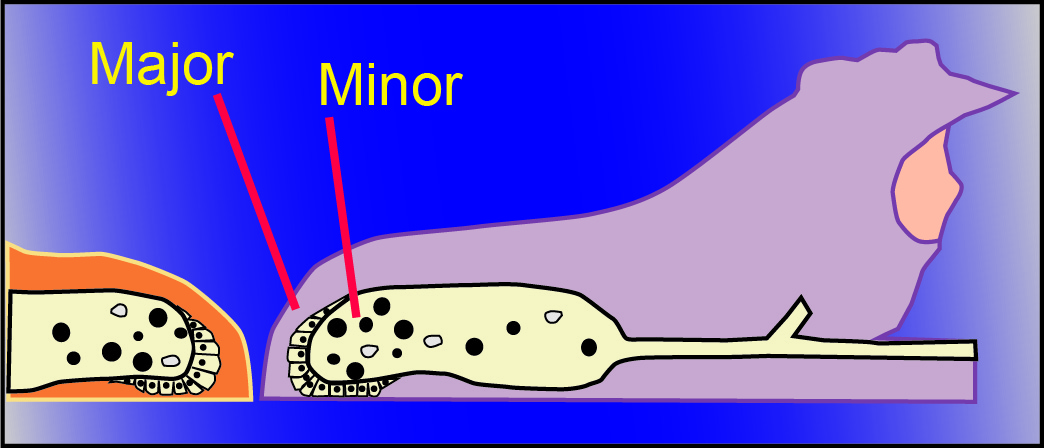
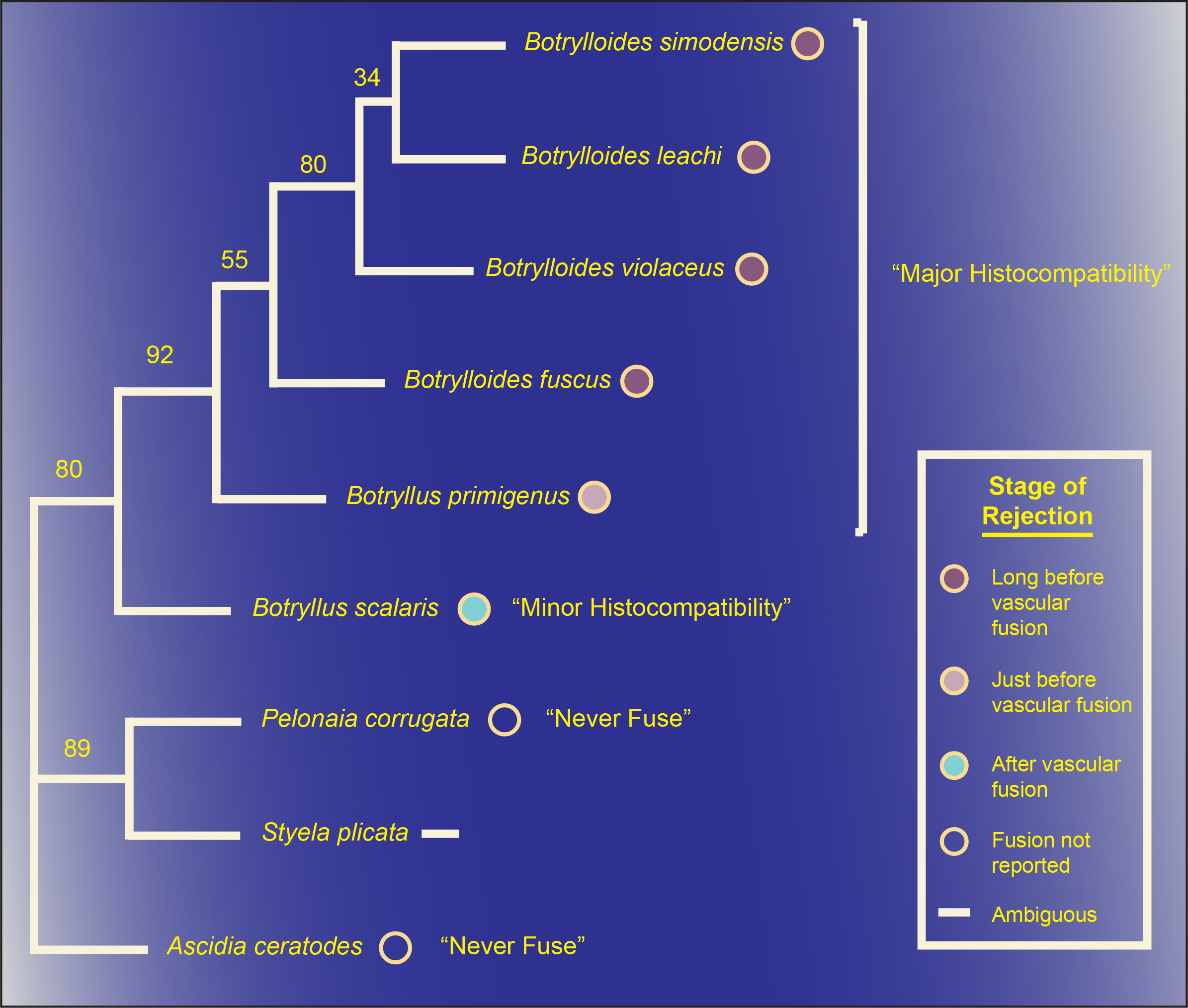
Association theory resolves a number of anomalies of including (1) why genetic kin recognition cues are evolutionarily stable, (2) why cheaters are rarely found in nature, (3) why some organisms associate preferentially with kin despite lacking altruistic behavior, and (4) why some organisms associate indiscriminately with both kin and nonkin.